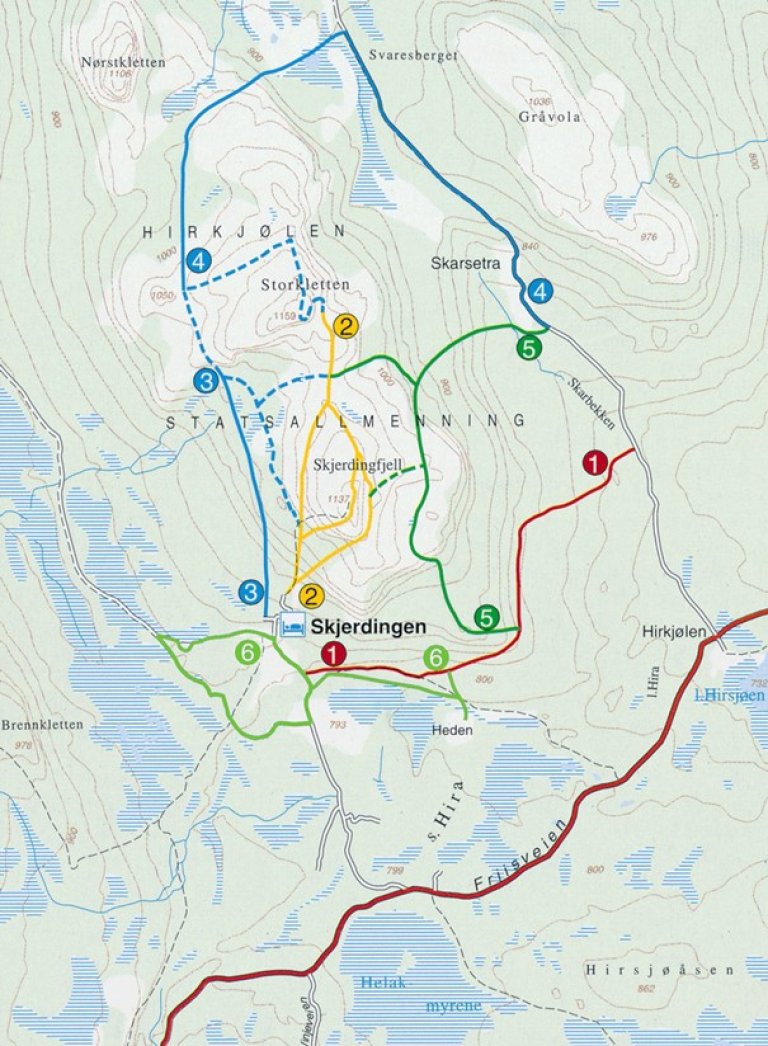
Hirkjølen demonstrasjonsområde har et tilbud til de fleste som liker friluftsliv og har interesse for geologi, planter, dyr og kulturminner. Det er seks naturstier av ulik lengde og med god variasjon innen temaer. Stiene er godt merket og utgjør til sammen 35 km. Deler av forskjellige stier er koblet sammen med tverrforbindelser slik at turen kan få en lengde og et innhold som passer deg.
Hirkjølen demonstrasjons- og forsøksområde ligger i Ringebu kommune ved Friisveien mellom Atna i Østerdalen og Ringebu i Gudbrandsdalen. Avstanden fra Oslo er 260 km, fra Ringebu 40 km og fra Atna 18 km.
Ved Skjerdingen Høyfjellshotell står en tavle med kart over stiene. Her finnes ellers en utstilling og stifoldere. Det er overnattingsmuligheter ved Hyggehotellet Skjerdingen tlf 62 69 99 90. Mer info om hotellet finner du på skjerdingen.com
Avstander:
Oslo 260 km - Trondheim 250 km - Lillehammer 100 km (Sommer) - Lillehammer 140 km (Vinter) - Elverum 130 km - Bjorli 210 km - Hamar 160 km - Atna 18 km - Ringebu 40 km (Sommer) - Ringebu 85 km (Vinter)
Elg i fangstgrop. Illustrasjon av Harriet Ask Kihle
Hirkjølen demonstrasjonsområde ligger i Ringebu kommune.
Området har seks ulike naturstier av ulik lengde og med fokus på ulike tema. På stiplakatene fokuserer vi på økologiske forhold i fjellskogen og menneskenes historiske bruk av denne. Temaer som klimaendringer, naturforvaltning, biologisk mangfold og fjellskogsbruk er belyst.
Foruten turgåere brukes Hirkjølen demonstrasjonsområde til undervisning, fagsamlinger innen skogbruk og forskning. Norges miljø- og biovitenskapelige universitet (NMBU) bruker området regelmessig til undervisning innen skogfag, jordbunnslære og geologi.
Også Evenstad (iNN) med sin nærhet til Hirkjølen, bruker området til forskning og undervisning. Flere mastergrader og forskningsprosjekter har blitt utført på Hirkjølen de siste årene, blant annet ved studenter fra Universitet i Bergen.
Skogavdelingen ved Evenstad på befaring i Hirkjølen demonstrasjonsområde. Foto: Per Holm Nygaard
Geologi
Under den kaledonske fjellkjedefoldningen ble forstenede grus- og sandavsetninger i dyphavet mot nordvest skjøvet innover grunnere havomåder ved Hirkjølen. Kjempestore steinflak ble stablet over hverandre og nådde antagelig opp til 1000 m over dagens topper. Disse fjellene er senere slitt ned gjennom mange istider, slik at bare rester etter skyvedekket ligger igjen. I bruddkantene mot sydøst finner du snitt gjennom disse gamle avsetningene.
Under Skjerdingfjell ligger et lag kalkstein med kalkholdig skifer over (sti 2).
Geologi. Foto: Jon Eivind Vollen
Planter
Sammen med klimatiske variasjoner gir dette grunnlag for et særlig stort utvalg av planter. Undersøkelser i 1930-årene viste 665 arter og underarter, og noen av disse er sjeldne i dette området. Senere undersøkelser har påvist flere spesialiserte mose-, lav- og sopparter i vernede deler av den gamle skogen i østlia.
Sydhellingene under Skjerdingfjell og Storkletten varmes raskt opp om våren. De tidligste artene (rødslidre og snømure) kan starte blomstringen alt ved månedsskiftet april/mai, mens snøen ennå ligger dyp i resten av området. Kalkholdig morene og kalksig gir næringsrik jord med rik blomstring gjennom sommeren. Du finner planter som er avhengige av kalk slik som reinrose, bergstarr og bakkesøte. Andre rike naturtyper finnes langs stinettet og på setervollene.
Dyr
Elg, rein og rådyr har faste stammer her, mens hjort av og til streifer innom. Alle de fire store rovdyrene ferdes av og til gjennom området.
Du kan treffe på eller se sportegn etter bever, rev, grevling, mår, hare, røyskatt, smågnagere, storfugl, orrfugl, lirype og en rekke arter av vade-, ande- og spurvefugler.
Vanlige rovfugler er kongeørn, fjellvåk, tårnfalk, dvergfalk og i perioder haukugle. Det er ørret i både bekker, elv og vann i nærområdet.
Ønsker du en mer dyptgående beskrivelse av plante- og dyrelivet, se nærmere på publikasjonen Hirkjølen - dyr og planter (lenke under "Publikasjoner" nederst på siden).
Kulturminner
Rikt beite og dyreliv, godt fiske, store forekomster av myrjern og etter hvert skogbruk og interessant natur har trukket folk til området gjennom mange tusen år. Torvanalyser viser at det var jegere og samlere her for nesten 9 000 år siden.
Husdyrbruk og korndyrking startet for 4 500 – 5 000 år siden, antagelig på de samme områder som det er beiter i dag. Elg ble fra gammelt av fanget i nær 100 fangstgroper som med sperregjerder stengte hele dalføret mellom Skjerdingfjell og Imsdalsvola. Sti 1 og 6 passerer to slike groper.
En overgang mellom Skjerdingfjell og Storkletten var stengt med «bogesteller», hvor det satt jegere med pil og bue og skjøt på passerende dyr (sti 2).
Myrmalm ble brukt til å fremstille jern fra starten av jernalderen. Jernholdig torv ble først brent med ved. Sti 1 passerer et blesteranlegg, og prosessen er vist ved et stort anlegg i sti 6.
I nyere tid begynte seterdriften i stor skala ved slutten av 1600-tallet og varte ut i 1950-årene. Det var helårsdrift på fire gårder ved Hirkjølen. Sti 1 og 6 passerer den ene av disse.
Bærekraftig skogbruk
Hirkjølen Forsøksområde er drevet som et senter for økologisk forskning i fjellskog og med omfattende registreringer av klima, jordbunn, flora og fauna siden 1930. Alle typer fjellskoghogst og foryngelsesmetoder er representert innen forsøksområdet. Fra gammel plukkhogst, bledningshogst, stripehogst, flatehogst, frøtrestillingshogst til dagens stedstilpassede fjellskogshogst hvor bærekraftig skogbruk er vektlagt. Kulturbestand av norske og utenlandske bartreslag anlagt i fjellbjørkeskog nær skoggrensen viser produksjonspotensialet og muligheter for karbonlagring i et endret klima.
Det er lagt vekt på å vise hvordan skogen kan forynges, og hvilke metoder som ikke lykkes i fjellnære områder, slik som storflatehogst og stripehogst. I dagens fjellskogbruk velges hogst- og foryngelsesmetoder som tar hensyn til klima, flora, fauna og friluftsliv i tillegg til tømmer produksjon.
Partier med gammel naturskog er vernet innenfor Skardseterlia naturreservat. Dette er samtidig viktige demonstrasjons- og forskningsområder, som er tilgjengelige gjennom sti 5. Skogbruk er ellers viktige tema i stiene 1 og 3.
Naturstiene ligger i Hirkjølen demonstrasjonsområde, og er et samarbeid mellom Statskog, Skogkurs og NIBIO.
Skogbruksledere fra Gudbrandsdalen på besøk i skogstien i 2025. Foto: Per Holm Nygaard
Ole Martin BollandsåsIngvild RekstadErik NæssetIngvald Røsberg
Sammendrag
The objectives of this study were (1) to develop models for estimation of total above-ground biomass, tree crown biomass and stem biomass of mountain birch (Betula pubescens spp. czerepanovii), and (2) to test the stability of the relationships between biomass and biophysical tree properties across geographical regions and tree size ranges. The models were developed using a mixed modelling approach accounting for the hierarchical structure of the data that originated from sample plots. Diameter at breast height, tree height, and the ratio between height and diameter were candidate explanatory variables, but only diameter was statistically significant (p0.05). The model fit values (pseudo-R 2) were 0.91, 0.60 and 0.85 for the three respective models. A substantial part of the model random errors could be attributed to between-plot variations. The conclusion related to objective (1) was that the models are well suited for biomass prediction of mountain birch in the mountain areas of southern Norway. Furthermore, models reported in previous research that had been calibrated on data from other regions were applied on the current data set. The results indicate that models calibrated for small trees produced predictions diverging from the observed values of the current data set. The differences between predicted and observed values also seem to vary along a site productivity gradient. Still, even though the differences between predicted and observed values using the different models varied quite a lot, the relationships were relatively stable within certain limits. The conclusion related to objective (2) was that biomass models can be applied outside the region for which they were developed, which in many cases is necessary because local models do not exist. However, the properties of the model development data related to tree size range and site productivity should be similar to those of the area for which predictions are being made.